![]()
![]()
![]()
![]()
![]()
![]()

![]()
![]()
|
|
|
Loss and recovery of wings in stick insects
MICHAEL F. WHITING*, SVEN BRADLER† & TAYLOR MAXWELL‡
The evolution of wings was the central adaptation allowing insects to escape
predators, exploit scattered resources, and disperse into new niches, resulting
in radiations into vast numbers of species1.
Despite the presumed evolutionary advantages associated with full-sized wings (macroptery),
nearly all pterygote (winged) orders have many partially winged (brachypterous)
or wingless (apterous) lineages, and some entire orders are secondarily wingless
(for example, fleas, lice, grylloblattids and mantophasmatids), with about 5% of
extant pterygote species being flightless2, 3.
Thousands of independent transitions from a winged form to winglessness have
occurred during the course of insect evolution; however, an evolutionary
reversal from a flightless to a volant form has never been demonstrated clearly
for any pterygote lineage. Such a reversal is considered highly unlikely because
complex interactions between nerves, muscles, sclerites and wing foils are
required to accommodate flight4. Here we show that
stick insects (order Phasmatodea) diversified as wingless insects and that wings
were derived secondarily, perhaps on many occasions. These results suggest that
wing developmental pathways are conserved in wingless phasmids, and that
're-evolution' of wings has had an unrecognized role in insect diversification.
Stick insects are large terrestrial insects that exhibit extreme forms of
morphological and behavioural crypsis as mimics of sticks and leaves. Phasmids
are chiefly arboreal insects, but a few of the more robust taxa (for example, Eurycantha,
Agathemera, Heteropteryginae) occur primarily near the ground, in the
leaf litter. The monophyly of Phasmatodea is supported by a series of
distinctive morphological characters including prothoracic repellant glands,
absence of mitochondria in spermatozoa, and male vomer5, 6.
Phasmids belong among the basal winged insect orders (Polyneoptera), but their
sister group is unknown, although Caelifera (grasshoppers)7,
Orthoptera8, Dermaptera (earwigs)6,
Grylloblattoidea and Dermaptera9, Dictyoptera (mantids,
cockroaches and termites)10, and Embiidina (web
spinners)6, 11 are all
candidates. It is difficult to decipher the phylogenetic relationships within
Phasmatodea because of the convergent morphology associated with their
remarkable crypsis. Current classification is based on some dubious
morpholological characters12, and no formal
investigation of phasmid phylogeny has yet been published. Of the 3,000 described species of phasmids placed in 3 families and
approximately 500 genera, only 40% are fully winged, with the remainder being
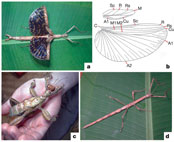
partially winged or wingless. In fully winged phasmids, the front wing is
reduced and thickened, and the hind wing is enlarged with an extensive anal
region (Fig.
1a). Vein homology is readily assessed between phasmids and other insect
groups, and analysis indicates that phasmid wings are homologous to those of
other insects (Fig.
1b). Partially winged phasmids have reduced hindwings with a small anal
region, and individuals are not capable of sustained flight (Fig.
1c). In wingless phasmids, no wing remnants are present (Fig.
1d). The extent of wing development of male phasmids is never less than
females: if the male is wingless, the female is always wingless; if the male is
partially winged, the female is partially winged or wingless, but never fully
winged; if the male is fully winged, the female may be fully winged, partially
winged or wingless.
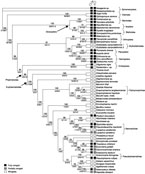
DNA sequence data were used to place the Phasmatodea among the polyneopterous insect orders and to estimate phylogenetic relationships for major stick insect lineages. The entire regions of 18S ribosomal DNA (18S rDNA; about 1,900 base pairs (bp)), 28S rDNA (2,250 bp), and a portion of histone 3 (H3, 372 bp) were sequenced from 22 outgroup and 37 ingroup taxa representing all Polyneoptera and 14 of the 19 recognized phasmid subfamilies. Trees were reconstructed through optimization alignment and nodal support determined by nonparametric bootstrap and partitioned Bremer support (see Methods and Fig. 2). Partitioned Bremer supports indicate that 31% of the signal is provided by 18S, 62% by 28S and 7% by H3. Overall, H3 provides resolution at the shallow nodes, 18S at the deep nodes, and 28S throughout the entire topology. Most of the nodes are very well supported, with 44 out of 56 nodes with bootstrap values of 95 or higher and 52 out of 56 nodes with a total Bremer support of 5 or greater. These data support the monophyly of each insect order, the basal placement of Timematidae, and the monophyly of the groups Euphasmatodea, Pachymorphinae, Necrosciinae, Pseudophasmatinae and Lanceocercata, in agreement with other studies5, 6, 8, 13. Analyses using maximum parsimony, maximum likelihood, and bayesian methods result in a topology that is highly congruent with the optimization alignment topology (see Supplementary Information).
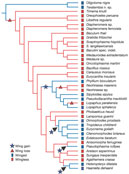
The most parsimonious reconstruction (MPR) of the states 'winged' and 'wingless' unambiguously places the ancestral state for phasmids as wingless, with wings derived on four (ACCTRAN optimization) or five (DELTRAN optimization) occasions (Fig. 3). Forcing the ancestral state as winged requires 13 steps in males and 14 steps in females, and is the MPR only when wing gain is weighted six times wing loss. Similar results are found when winged is divided into 'fully winged' and 'partially winged' and the states are treated as unordered. Wing states were also mapped by means of likelihood methods on both the bayesian and likelihood trees, to take into account branch lengths in determining probabilities of ancestral states14. When the rate of wing gain is set equal to that of wing loss, the phasmid ancestor is reconstructed as wingless (P < 0.001), with four independent wing gains in more derived stick insect lineages. When the ratio of the rate of wing loss over wing gain ranges from 1 to 1,400, there is a >95% probability that the ancestral phasmid was wingless.
These results support the hypothesis that the ancestral condition in Phasmatodea is wingless, that the first six basal phasmid lineages are entirely wingless, and that fully developed wings were derived later in phasmid evolution, on as many as four occasions. Clearly, the presence of wings is a very plastic feature in phasmids, with congeneric species (for example, Lopaphus) exhibiting both partially winged and wingless states. One of the correlates with winglessness in insects is increased female fecundity2, 3, and as phasmids scatter specially modified eggs individually rather than concentrating them in large numbers similar to their sister taxon, there may have been a selective advantage early in phasmid evolution to shift to winglessness to facilitate fecundity and increased crypsis. The detailed homology in wing features shared among phasmids and other insects suggests that wings did not re-evolve de novo in phasmids, but are rather a re-expression of the basic insect wing which was lost in ancestral stick insects. Entomologists have long assumed that re-evolution of wings in apterous lineages was impossible, because functional wings require complex interactions among multiple structures, and the associated genes would be free to accumulate mutations in wingless lineages, effectively blocking the path for any future wing reacquisition. However, this assumption requires that developmental pathways for wing formation are largely independent of pathways required for development of other structures. For instance, in Drosophila and other insects, leg and wing imaginal discs have a common origin from a single group of cells and the developmental pathway for wing determination has been largely co-opted (recruited) from the pathway required for limb formation15, 16. Therefore it is not surprising that the basic genetic instructions for wing formation are conserved in wingless insects, because similar instructions are required to form legs, and probably other critical structures16. Studies of flight motor patterns in flying and non-flying phasmids indicate that the non-flying phasmids have retained the neural structures and basic functional circuitry required for flight, as indicated by flight-specific neural activity in thoracic muscles17, demonstrating that the loss of wings does not correlate with the loss of flight musculature and innervation. Wing development depends on multiple gene systems, transcription factors, secreted proteins, and receptors15, and mutations in any one of these factors may lead to winglessness. Given the multitude of factors involved in wing formation, it seems probable that the specific cause for winglessness will differ from lineage to lineage, but that the basic blueprint for wing formation can remain largely intact, even over large evolutionary time periods. Re-evolving complex structures from the same basic building blocks may be a more general trend in evolution than previously recognized. For example, it is plausible that all modern animals with eyes evolved from a common ancestor that possessed a primitive image-forming organ18, controlled by the master gene Pax-6. Our results support the hypothesis that the developmental pathway for wing formation evolved only once in insect diversification, but that wings evolved many times by silencing and re-expressing this pathway in different lineages during insect evolution. Thus, wing loss does not seem to be an evolutionary dead end, and the ability to regain a wing over evolutionary time means that lineages have the adaptive advantages of being both winged and wingless. The transition from wingless to flying forms may be a common theme in insect evolution, as it has been suspected within one genus of water striders19, and may occur in cockroaches and Hemiptera, which also exhibit a wide diversity of wing forms. To our knowledge, this is the first example of a complex feature being lost and later recovered in an evolutionary lineage, and it is possible that the reacquisition of complex features may have an important role in evolutionary diversification. Methods Alignment and phylogenetic analysis Alignment of histone 3 was based
on conservation of amino acid reading frame. A gross alignment was performed on
the ribosomal genes by manually aligning the conserved domains across all taxa
in Sequencher 3.1.1. Each conserved domain, and the variable regions between the
conserved domains, underwent optimization alignment using the computer program
POY, run in parallel mode across an IBM SP2 Supercomputer; this is the first
implementation, to our knowledge, of this program on a supercomputer. Ribosomal
expansion regions were excluded from outgroups but aligned for ingroups as
described elsewhere22. Sixteen parameter sets
(gap/change ratio 1–4; transition/transversion ratio 1–4) were applied
across all data partitions (18S rDNA, 28S rDNA, H3, and total). The parameter
combination with gaps, transitions and transversions weighted equally minimized
incongruence among data partitions (incongruence length difference (ILD) metric
= 0.01722), and was selected as the best-justified parameter set for
optimization alignment23. Topologies from
parameter combination sets adjacent to the optimal combination produced similar
phylogenetic results. Optimal parameters produced a single topology (length =
9,057, consistency index = 0.508, retention index = 0.618), and the implied
alignment from this topology was used to calculate bootstrap and partitioned
Bremer support values. Bootstrap values were calculated in PAUP*4.0b10 (ref. 24)
with 10,000 replicates, 5 random additions per replicate, and TBR branch
swapping. Partitioned Bremer support values were calculated using TreeRot25.
The incongruence length difference test26
indicated insufficient evidence to reject the hypothesis of data set congruence
(P = 0.892), therefore individual data sets were combined into a total
evidence analysis. The states 'fully winged', 'partially winged' and 'wingless'
were treated as unordered characters and mapped for males and females by means
of parsimony using MacClade 4.0 (ref. 27). For the ingroup,
the MPR required ten steps (males) and nine steps (females), the ancestral
phasmid state was reconstructed unambiguously as wingless, and wings were
derived four times unambiguously. Constraining the ancestral phasmid state to
fully winged (by forcing state changes as irreversible) required 28 steps for
males and 30 steps for females, and is the MPR when wing gain is 11 times wing
loss. Fully winged and partially winged were combined into winged and the MPR
required 7 steps (males and females), the ancestral state was reconstructed as
wingless, and wings were unambiguously derived on four occasions. Mapping of
characters by means of likelihood was performed using the program Discrete28
on topologies generated by means of standard likelihood and bayesian methods
(see Supplementary
Information). Wings were mapped for males and females (character 1 and 2)
with two states: wings present or absent. Character states were mapped on the
maximum likelihood topology and the bayesian topology with the rate of wing loss
set to a value of 1, and the rate of wing gain set to 20 individual values,
spanning the interval 1 to 0.00001. Character states were also mapped on a set (n
= 30) of bayesian trees29, and the mean
probability that the ancestor to Euphasmatodea was wingless is 0.9996 Supplementary information accompanies this paper. Received 29 May 2002;
Acknowledgements. We thank E. Tilgner for providing some specimens; A. Whiting and L. Harmon for assistance in data analysis; and J. Cherry, M. Gruwell, M. Terry, H. Ogden, J. Robertson and K. Jarvis for generating sequence data. Analyses were performed in the Fulton Supercomputer Center, Brigham Young University, and parallel software implementation was performed by M. Clement and Q. Snell. DNA sequences are deposited in GenBank under accession numbers AY121129–AY121186 and AY125216–AY125326.
|
Send mail to mel@tungate.com with
questions or comments about this web site.
|